

和缓艾美耳球虫是一种致病性较弱的鸡艾美耳球虫,主要寄生在回肠、盲肠和直肠,其在英国、美国、澳大利亚、约旦、捷克和瑞典等多个国家均有报道,在我国多个省市也有发现报道 [1] ,且部分地区存在较高的流行,如成都 [2] 、山东 [3] 、安徽某些地区 [4] 、江苏徐州 [5] 等地区均有和缓艾美耳球虫是鸡球虫病优势种之一的报道。赵世明等人 [3] 对山东省 50 家养殖条件差且出现亚临床球虫病的肉鸡场调查,显示和缓艾美耳球虫感染率达 60%。和缓艾美耳球虫低剂量感染后无明显可见的病变,仅在高剂量感染后表现为生长缓慢,产蛋减少,饲料转化率降低,有时可见水样稀粪,严重可致死亡 [1] ,因而在早期的鸡球虫病研究中未受重视,关于它的研究报道较为少见。自 Jeffers [6] 1975 年首次报道柔嫩艾美耳球虫株早熟系选育技术以来,该技术已被证明为适用于所有鸡艾美耳球虫种 / 株致弱的可靠和高效方法。笔者在分离和纯化出和缓艾美耳球虫保定株的基础上,对其进行了早熟选育及早熟系生物学特性研究,以期对制备含有和缓艾美耳球虫早熟系的鸡球虫病疫苗提供前期研究基础。
1、材料与方法
1.1 实验动物及其饲养
从济南斯帕法斯家禽有限公司购进 3~7 日龄 SPF 鸡,分别隔离饲 养于隔离器中,饲喂无抗球虫药物的全价饲料,用前经高压蒸汽灭菌, 供饮用水为纯化水。试验前,检查粪便无球虫卵囊污染。
1.2 实验虫株
和缓艾美耳球虫保定株(MiBD 株)由佛山市正典生物技术有限公 司分离、鉴定和保存。
1.3 早熟选育
用和缓艾美耳球虫保定株孢子化卵囊经口感染 30 只 7 日龄 SPF 鸡。从所接种球虫潜隐期的前 2 h 开始每隔 1 h 进行饱和盐水漂浮法粪检,当检测到成熟卵囊,并预计能收获下次早熟选育所需卵囊数的 2 倍卵囊时,全部剖杀鸡只,收取粪便,并把最早发现卵囊的时间记录为该代次卵囊的潜隐期。重复上述传代过程,直到潜隐期缩短到一定程度为止。
1.4 早熟系生物学特性测定
1.4.1 早熟系潜隐期稳定性测定
早熟选育至一定代次后,对所收获早熟系的潜隐期进行稳定性测定。取 7 日龄 SPF 鸡 10只,每只口服感染被检早熟系孢子化卵囊 5×103 个。监测子代卵囊的潜隐期,收集其卵囊排泄高峰期排出的粪便,粪便中的卵囊经分离和孢子化后标记代次,将所收获的卵囊重复上述传代过程,共传代 5 代次。
1.4.2 排卵囊规律测定
取体况相近的 14 日龄 SPF 鸡 30 只,随机分为 3 组,每组 10 只。各组分别经口感染纯种和缓艾美耳球虫保定株早熟系孢子化卵囊 5×102 个 / 鸡、1×103 个 / 鸡和 1.5×103 个 / 鸡。在潜隐期(82 h)前 1 天开始每隔 1 h 用饱和盐水漂浮法进行粪检,检测排卵开始时间。卵囊开始排出后,每隔 24 h 收集粪便 1 次,计算每克粪便卵囊数(OPG),直至 OPG 降至较低水平,同时计算其平均每羽鸡卵囊产量。通过对每日排卵囊量的统计获得和缓艾美耳球虫保定株早熟系的排卵囊规律,包括排卵囊持续时间和排卵囊高峰期。相关计算公式:
OPG=a×100;a 为麦氏计数板显微镜下计得的卵囊数平均每只鸡卵囊产量=OPG×粪重÷鸡数量。
1.4.3 早熟系致病力测定
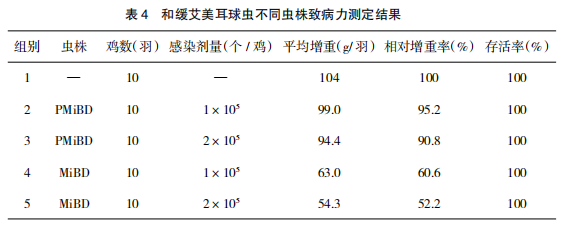
取 7 日龄 SPF 鸡 50 羽,随机分成 5 组,每组 10 羽。感染前每组鸡只进行称重,并进行以下处理:第 1 组为空白对照组,第 2、3 组为感染早熟系,第 4、5 组感染亲本株(MiBD 株),感染剂量分别为 1×105 、2×105 个孢子化卵囊。感染后,观察各组鸡只精神状态、饮食欲,4 天后观察鸡只粪便情况,第 6 天进行称重。统计死亡率并计算各组平均增重和相对增重率。
1.4.4 早熟系免疫原性测定
取 3 日龄 SPF 鸡 50 只,分成 5 组,每组10 只,其中第 1、2、3 组为免疫攻毒组,每鸡分别口服接种 50、100、150 个早熟系孢子化卵囊;第 4 组为不免疫攻毒阳性对照组;第 5 组为不免疫不攻毒阴性对照组。将 5 组鸡分别在相同条件下隔离饲养(垫料上饲养)。免疫后第21 日,阳性对照组、免疫攻毒组鸡只分别口服攻击和缓艾美耳球虫保定株(MiBD 株)强毒孢子化卵囊各 1×105 个,观察 5~6 日,称量、统计各组鸡体重并计算相对增重,分别收集攻毒后 120~144 h 阳性对照组和免疫攻毒组鸡粪便并计数克粪便卵囊数(OPG),计算免疫攻毒组相对于阳性对照组的卵囊减少率。相关计算公式:
卵囊减少率(%)=(阳性对照组 OPG-免疫攻毒组 OPG)÷阳性对照组 OPG×100%
2、结果
2.1 早熟系选育结果
2.1 早熟系选育结果
经过 11 代的早熟系选育,和缓艾美耳球虫保定株(MiBD 株)潜隐期由 97 h 降至 82 h,将此早熟系命名为和缓艾美耳球虫保定株早熟系,代号为 PMiBD 株。具体选育过程和结果见表 1 和图 1。
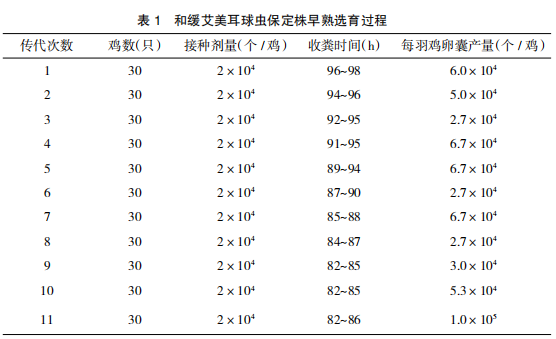
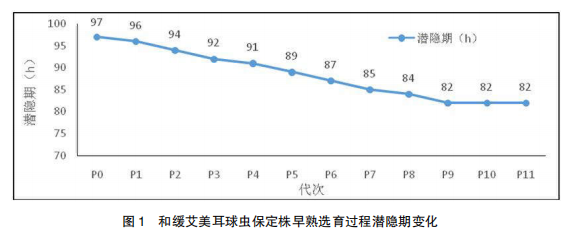
2.2 和缓艾美耳球虫保定株早熟系(PMiBD株)潜隐期稳定性测定结果
潜隐期为 82 h 的和缓艾美耳球虫保定株早熟系(PMiBD 株),经 5 代次无早熟选择压力传代,其潜隐期仍为 82 h。
2.3 排卵囊规律测定结果
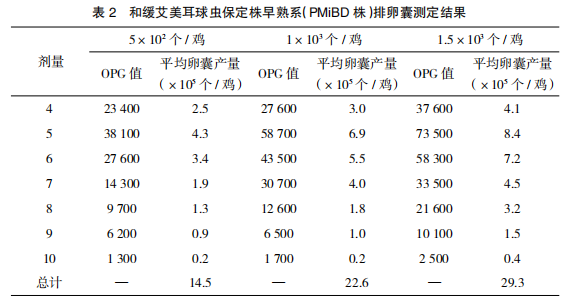
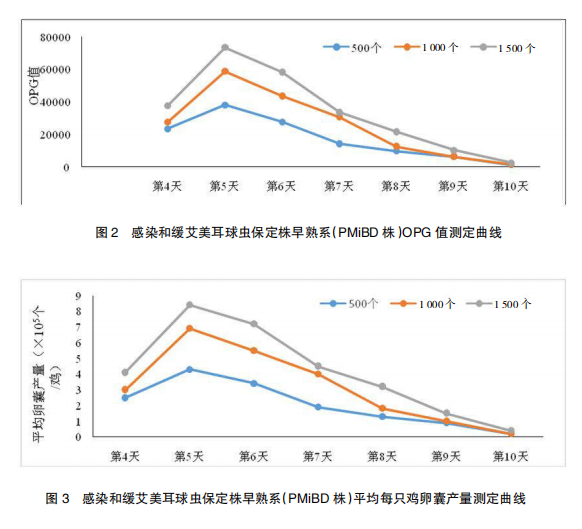
感 染 和 缓 艾 美 耳 球 虫 保 定 株 早 熟 系(PMiBD 株)孢子化卵囊后 82 h 开始有卵囊排出,从感染后第 4~10 天检测粪便 OPG 值,结果表明:感染和缓艾美耳球虫保定株早熟系(PMiBD 株)孢子化卵囊后 OPG 值和平均每羽鸡卵囊产量均随着感染剂量的增大而增大,且感染后第 5 天为排卵高峰期,随后逐步降低,第 10 天 OPG 值已明显下降至 3.0×103 以下。感染后第 4~6 日的排卵囊量占卵囊总产量的70.3%(500 个 / 鸡)、68.1%(1 000 个 / 鸡)和67.2%(1 500 个 / 鸡)。结果见表 2、图 2~3。2.4 和缓艾美耳球虫保定株早熟系(PMiBD株)致病力测定结果
和缓艾美耳球虫保定株早熟系(PMiBD株)高剂量感染后鸡只精神状态、饮食欲无明显异常,粪便形态正常或仅有少量水样粪便、饲料粪;亲本株(MiBD 株)感染组出现该球虫引起的明显临床症状:精神状态沉郁、饮食欲明显降低或不食、水样粪便或饲料粪。另外,同剂量感染的早熟株和亲本株感染组相比,早熟株感染组鸡只感染后相对增重明显高于亲本株感染组(详见表 4),说明和缓艾美耳球虫保定株早熟(PMiBD 株)的致病力显著低于其亲本株。2.5 和缓艾美耳球虫保定株早熟系(PMiBD株)免疫原性测定结果
和缓艾美耳球虫保定株早熟系(PMiBD)3个不同免疫剂量组试验鸡,免疫后攻毒,阳性对照组的相对增重率低于 80%,其中免疫剂量 50个卵囊 / 鸡免疫攻毒Ⅰ组相对增重率为 71.6%,仅高于阳性对照组 8.1%;免疫剂量 100、150 个卵囊 / 鸡免疫攻毒Ⅱ组和Ⅲ组的相对增重高于其阳性组 20%以上。与阳性组鸡粪便 OPG 相比,免疫剂量 50、100、150 个卵囊 / 鸡的卵囊减少率均不低于 80%,说明该早熟系具有良好的免疫原性(详见表 5)。

4、讨论与分析
目前关于和缓艾美耳球虫的研究报道较为少有,对其早熟系的研究报道则更为鲜有,仅有几篇报道。McDonald V 等人 [7] 1983 年报道了 1株和缓艾美耳球虫豪顿株经过 10 代早熟选育潜隐期从 93 h 缩短至 73 h,随后对其繁殖力、免疫原性进行了研究,发现该早熟株繁殖力较亲本株弱,免疫原性良好,且若干代繁殖后无毒力返强 [8,9] 。索勋 [10] 2005 年报道了 1 株和缓艾美耳球虫涿州株经过 15 代早熟选育后潜隐期从96 h 缩短至 77 h,且该早熟株致病性与繁殖力均较母株弱,但保留了母株的免疫原性,经 10代非早熟选择性传代后,其潜隐期、繁殖力和致病性均稳定。王娟等人 [11] 2016 年报道了 1 株和缓艾美耳球虫巴彦淖尔株经过 15 代早熟选育潜隐期从 96 h 缩短至 77 h,该早熟株具有良好的遗传稳定性和免疫原性,且致病性和产卵囊量降低。而笔者分离得到的和缓艾美耳球虫保定株在经过 11 代早熟选育后潜隐期从 97 h 缩短至 82 h,该早熟系的致病性均较亲本株弱,但免疫原性良好,每羽鸡免疫 100 个以上卵囊即可较好抵抗 1×105 个亲本株的感染,相对增重率达 80.0%以上和卵囊减少率达 90.0%以上,这些发现均与前人报道的结果相似。早熟选育技术的成熟运用可以获得弱毒且免疫原性良好的鸡艾美耳球虫虫株,为各种鸡球虫病弱毒活疫苗的研发提供了有效技术手段。
5、结论
经过对和缓艾美耳球虫保定株连续 11 代的早熟选育,我们获得了潜隐期由 97 h 缩短至82 h 的早熟系。该早熟系的潜隐期稳定,感染后4~10 天为排卵期,第 5 天为高峰,致病力明显低于其亲本株,且具有良好的免疫原性。
参考文献
[1] 赵 爱 云 , 段 嘉 树 . 和 缓 艾 美 耳 球 虫 (Eimeriamitis)研究进展[J]. 中国兽医寄生虫病, 2006(02):28-33.
[2] 杨发龙, 蒋锡仕, 李必富, 等. 成都地区鸡球 虫种类的初步调查[J]. 四川畜牧兽医, 2001(04): 24-29.
[3] 赵世明, 孙希萌, 索勋. 山东省 50 家肉鸡场 和缓艾美耳球虫流行病学调查 [J]. 中国兽医杂 志, 2009(03):31-33.
[4] 肖寒, 李培英, 王俊生, 等. 安徽省部分地区 鸡球虫种类及感染情况调查 [J]. 中国兽医寄生 虫病, 2008(02):18-22.
[5] 丁文卫, 芮艺, 夏冰, 等. 徐州地区 16 个小 型鸡场球虫种类的调查 [J]. 畜牧与兽医, 2010 (11):69-72.
[6] Jeffers T K. Attenuation of Eimeriatenella through selection for precociousness [J]. J Parasitol, 1975,61(6):1083-1090.
[7] McDonald V, Ballingall S. Attenuation of Eimeriamivati (= mitis) by selection for precocious development [J]. Parasitology, 1983,86 (Pt 3): 371-379.
[8] McDonald V, Shirley M W. Eimeriamitis: a comparison of the endogenous developmental stages of a line selected for early maturation and the parent strain[J]. Parasitology, 1984,88 ( Pt 1):37-44.
[9] McDonald V, Shirley M W, Chapman H D. Attenuation of Eimeria species: further characterisation of two lines of Eimeriamitis[J]. Res Vet Sci, 1985,39(3):328-332.
[10] 索勋. 和缓艾美耳球虫早熟系选育及其生 物学特性研究 [J]. 畜牧兽医学报, 2005,36(6): 602-605.
[11] 王娟, 刘永清, 高永刚, 等. 鸡和缓艾美耳球 虫早熟虫株选育及其生物学特性研究 [J]. 畜牧 与饲料科学, 2016(09):15-18.
作者:刘丽丹1王文莉1伍存1曾莉1唐建红1梁正鹏1位晓莉1翁亚彪1,2(1 广东省佛山市正典生物技术有限公司 , 广 东 佛 山528138,2 华南农业大学兽医学院,广州510642)
作者简介:刘丽丹(1983-),女,广东海丰人,硕士,中级兽医师,主 要从事动物寄生虫病及其防控研究。
通讯作者:翁亚彪(1963-),男,汉族,广东化州人,硕士,副教授,主 要从事球虫免疫技术和动物寄生虫病临床防控技术研究,E-mail: ybweng@scau.edu.cn。
基金项目:国家重点研发计划(2016YFD0501303)。
责任编辑:翁亚彪





